商机详情 -
河北脂质体载药mRNA
利用设计的脂质,他们发现由1,2-二油醇-3-二甲基氨基-丙烷(DODMA)阳离子脂质组成的核酸脂质颗粒在小鼠和食蟹猴中分别以0.01mg/kg和0.3mg/kg的剂量包封siRNA时表现出基因沉默作用。**近的一项构效关系研究表明,脂质结构的细微差异可能导致转染效率的明显差异。作者设计并合成了1,4,7,10-四氮杂环十二烷环基和含咪唑的阳离子脂质,它们具有不同的疏水区域(例如,分别为胆固醇和双薯蓣皂苷配基)。结果表明,这两种阳离子脂质在HEK293细胞中诱导有效的基因转染。脂质体各组分对核酸递送效率的影响。河北脂质体载药mRNA

脂质体用于抑菌的***除了脂质体**药物,第二大类脂质体药物是杀菌剂。两性霉素B是一种广谱多烯***,已经在医学上使用了几十年,被认为是***侵袭性******的金标准。它以细胞膜为靶点,与含胆固醇的哺乳动物细胞膜相比,对***细胞典型的含麦角甾醇膜表现出更高的亲和力。两性霉素B虽然具有很高的抗***活性,但也有严重的副作用,尤其是肾毒性。它是两亲性的,具有复杂的自关联行为,不同类型的聚集体表现出不同的溶解度和毒性;聚集状态也与药物疗效相关。因此,控制药物的聚集状态可以增强其***效果并降低其毒性。这种聚集控制是通过脂质纳米配方实现的。几种基于脂质的纳米颗粒制剂。两性霉素B已被开发出来,表现出良好的药代动力学特征,并***减少该药物的副作用。长沙脂质体载药荧光由于在巨噬细胞上发现了甘露糖受体, 因此甘露糖已被用于修饰阳离子脂质体以靶向巨噬细胞递送。
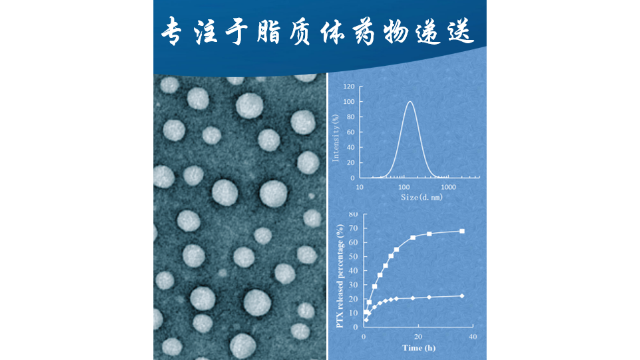
microRNA脂质体
microRNA是真核细胞中发现的短(约22mer)非编码RNA,通过结合互补的mRNA序列发挥生物调节剂的作用。miRNA以初级miRNA的形式从其编码的核基因转录,其长度为数百个核苷酸。RNaseIII酶,Drosha,将初级miRNA加工成pre-miRNA(长度为70个核苷酸),携带一个特征的发夹环。然后pre-miRNA移动到细胞质中,在那里RNaseIII酶Dicer产生成熟的miRNA和乘客链。***,成熟的miRNA被整合到RNAi诱导的沉默复合体中,以降解它们的靶mRNA。由DOTMA、胆固醇和vitaminETPGS1k琥珀酸盐组成的阳离子脂质体被证明可以有效递送pre-miRNA-133b,导致A549非小肺*细胞中成熟miRNA-133b的表达比对照组细胞增加2.3倍,Mcl-1蛋白的表达减少1.8倍。经尾静脉注射含有pre-miRNA-133b的阳离子脂质体(1.5mg/kg)的ICR小鼠肺组织中成熟miRNA-133b的表达比接受含有紊乱的pre-mirna的阳离子脂质体的小鼠高52倍。
阳离子脂质体的递送的优势各种基于核酸的分子已被研究作为下一代***药物,包括质DNA,反义寡脱氧核苷酸(as-odn),小干扰RNA(siRNA)和微RNA(miRNA)。这些分子共享各种物理化学性质,比化学药物更大,并且携带高度负电荷,限制了它们的细胞递送。因此,核酸疗法要想取得成功,就必须在识别合适的靶蛋白的同时,开发新的递送系统。质粒DNA是**早被认为用于***目的的核酸之一,长期以来一直在基因***的背景下进行研究。在此背景下,研究人员已经将重点放在病毒载体上,它可以赋予高转染效率和基因表达的连续调节。然而,Gelsinger在使用腺病毒载体的临床试验中不幸死亡,让人们意识到病毒材料可能并非完全安全。此外,病毒载体作为候选药物有几个缺点,例如需要为每个靶分子设计载体的不便,以及缺乏关于细胞表达剂量依赖性的知识。脂质体制备方法:二次乳化法。

适用于脂质体载药的荧光染料一些常用于标记脂质体的荧光染料包括:1.DiO(DiOC18(3)):DiO是一种疏水性的荧光染料,可以插入到脂质双层中,用于标记脂质体的膜。它在绿色波长下发出荧光。2.DiI(DiIC18(3)):类似于DiO,DiI也是一种疏水性的荧光染料,可以插入到脂质双层中。它在红色波长下发出荧光。3.RhodaminePE:RhodaminePE是一种红色荧光染料,常用于标记脂质体的表面。它具有良好的荧光稳定性和光学性能。4.NBD(Nitrobenzoxadiazole)衍生物:NBD衍生物是一类疏水性荧光染料,常用于标记脂质体内部的脂质分子。它们在蓝色至绿色波长下发出荧光。5.BODIPY(4,4-Difluoro-4-bora-3a,4a-diaza-s-indacene)衍生物:BODIPY衍生物也是一类常用的脂质体标记染料,它们具有较强的荧光信号和良好的化学稳定性。这些荧光染料可以根据需要选择不同的激发波长和发射波长,以满足实验的要求,并且它们通常与脂质体中的脂质相容,不会对脂质体的结构和性质产生***影响。脂质体疫苗可以机器体内的免疫应答。辽宁肝脏靶向脂质体载药
聚乙二醇在免疫脂质体中起到了重要作用。河北脂质体载药mRNA
脂质体疫苗通常在已知疫苗中使用纯化抗原或减毒病原体作为免疫原。然而,长期的免疫反应可能不会由纯化抗原诱导,甚至有时根本不会诱导反应。另一方面,减毒疫苗可以在免疫的患者中产生应答。然而,递送包裹在脂质体内的抗原可诱导长期应答,这在某些抗原的直接免疫中没有观察到。研究表明,恶性细胞的细胞膜可以形成包封潜在抗原的脂质体。文献报道了包封在脂质体中的肽作为**疫苗的***应用能力。有研究评估了BLP25(一个含有合成人MUC1肽的25个氨基酸序列)作为**疫苗的能力。用二硬脂酰磷脂酰胆碱、胆固醇和二肉豆醇酰磷脂酰甘油(摩尔比3:1:25)中含有的单磷脂酰脂A(1%w/w)制备脂质体,然后与脂偶联和非偶联的MUC1肽结合。C57BL/6小鼠免疫分别采用肽相关脂质体、肽与无肽脂质体混合、脂肽单独免疫。结果表明,脂质体制剂对免疫应答有深远的影响。与物理相关的脂质体观察到强烈的免疫反应(抗原特异性t细胞细胞反应),而与肽混合的无肽脂质体或单独的脂肽则没有。体液免疫反应受到关联性质的***影响,这可以通过表面暴露的肽脂质体诱导muc1特异性抗体来证明。因此可以通过调整脂质体药物传递系统来诱导优先细胞反应这提出了一个假设即不同的脂质体配方刺激不同的免疫途径。河北脂质体载药mRNA